INTEGRATED NEUROPSYCHOPHYSIOLOGICAL THEORY ABOUT LATERALIZATION AND CODING OF CEREBRAL HEMISPHERES
Michele TRIMARCHI
Published on 'Brain and science integration', 1983,1,17-37 - 'International Journal of Psychophysiology',1984,4,253 - Introduced at the First International Meeting on 'Science - Education - Evolution' Roma, 1982, and, afterwards, at XXXV World Medical Assembly, Venice, Cini Foundation, 24-28 Ottobre and at III International Congress of IOP - International Organization of Psychophysiology, Vienna University, July 11th 1986
PREMISE
The present theory integrates the several researches performed in neurophysiological field inherent in the interactions and
interrelationships between the two cerebral hemispheres. Therefore I think it’s useless reporting the long bibliography that brought me to formulate such a theory, as I should quote
with deserves and worth not only the most famous researchers like Roger W. Sperry and his equipe, but many others, less famous but as much worthy, who contributed to such a formulation. I
leave therefore to the researcher the analysis of this theory, informing that for a scientific proof a lab is not particularly required, as we can act directly on the person in front of
us, and on ourselves through stimulus and images, verbal or other kind of sounds we use everyday in our interactions. By analyzing the responses evoked and their times, we can have
a clear vision of this experimentation. A lot of demonstrative experiments will be published on the next issues of this magazine and others.
In every phenomenon the physical-chemical-biological principles rule the morphing of the
interactions of functional parameters, and the consequent expressions determine the reality of the phenomenon instant by instant.
The effects generated by filogenetical and ontogenetical etch on next transformations, in a
chain bringing together diversities and differentiations of all natural elements and every structure, except man potentially, binded by precise laws, can’t help following the line and the
direction of such an evolution.
The energetic morph-physiological reality of mankind locates him on a completely different base
than every other structure, even those that palaeontology and zoology define as similar species; Man is the only capable to etch on physical-chemical-biological principles, so much to
alter the line and the direction of the natural development of phenomena and have the power to destroy the global ecosystem and consequently self-destroy. The self-destruct power
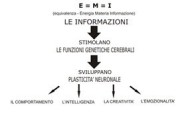
mankind actually owns is directly proportional to the non-knowledge of physical biological and energetic laws that rule us and determine our automatic and instinctive behaviour. First act
aiming to defuse such a power is the informational acquisition that every human being has a brain and every thought and action depend unambiguously on that, in relation to the synthesis
that takes place within the brain between the physiology of mechanisms and the learning of
information.
The evolution of atomic elements and the biological evolution may be integrated in a unique
evolutional process. In fact the evolution of atomic and molecular elements has never stopped and we can say that the long atomic experimentation, developed on the long path determined by
time and space, has brought to a continuously higher complexity with an order that partially still today escapes the researcher. Effectively all researchers know that the nervous
cell as we know it today is the result of a long biological evolution which started after and not together with the appearance of biological life on Earth. This kind of cell, that reaches
his top functional complexity in mankind, does not reproduce within its genetic nucleus, and has the same duration of the biological cycle of man: this is to demonstrate that it
represents our vital cycle. The several billions cells of our nervous system have to work in an environment being physiological to their functions and, in order to work, they have
to be physiologically stimulated. If we consider that every cell is different, this means they have different duties, therefore in order to work they need to have several billions
stimulus. How much do we really stimulate our cells? Are the stimulus physiological to their functions?
The repetitiveness of cultural models prevents them from working in a physiological manner on
the base of their diversity; this is why we use just a little of our brain.
This theory will allow to arrange at its best the culture and the life of a man in order to
eliminate the several problems that affect today all societies worldwide.
PHYSIOLOGY OF DUALISM OF MAN – THE TWO CEREBRAL HEMISPHERES
The anatomy of nervous system highlights the duality of the basic component elements functional to the fundamental duality of the brain, as
within it the cultural man and his story take place: the two telencephalic hemispheres.
Particularly, all of incoming sensorial stimulations have double driving channels, linked
through nervous ways to both of left and right hemisphere, so that every input is memorized independently in one and the other, evoking two different cortical responses.
If, from a morphological-anatomical point of view the cerebral hemispheres look uniformly
structured and differenced only by the mirror symmetry, their physiology highlights the fundamental differences that exist and determine the functional mechanisms, which are identified by
parameters which highlight the differences of coding and energy of neuronal cells and their connections.
One first differentiation is within the distribution of encephalic areas, which have different
functions in left or right hemispheres. A classic example is supplied by the centres of language and writing: although motor systems of vocalization have correspondence in both left and
right hemisphere, generally the ‘word’ has its site only in the left hemisphere.
Considering other aspects of physiology, a global and deeper difference between the two
hemispheres is defined by their different way to interact with outside environment, above all as to the responses to the stimulations coming from that.
Right and left hemisphere respond in a totally different way and time to
any
stimulus, as the energetic level and characteristic frequencies of interhemispherical biochemical stimulating functions are
different.
Through the duplicity of sensorial conditions, as above, the same informative message
follows different nervous ways and reaches the two hemispheres which elaborate and memorize it, giving a global worth of it which directly depends on the structure of the acquired
parameters of analysis. The physiology of left hemisphere is such to memorize information as cultural models, as what they have been until now, represented by the total of all
characteristics the physical information carries within, and makes this model an ‘absolute truth’ to be defended. The right hemisphere, instead, analyses all of the aspects of the
information and integrates them within, finding a common evolutional line that determines a spatial distribution sequence in no way limited by previous acquisitions.
By standing perceptively in front of an element, the two hemispheres select and code the
electro-magnetic information following different processes; e.g., through sensorial receptors of sight, they perceive the existence and the characteristics of an element: the
tree.
In the left hemisphere such a datum is identified instantly in a model and every revoking
function of the element ‘tree’ goes to join the specific tree model which has arrived to the hemisphere first. Definitively, the first memorization necessarily conditions next ones, as
these will always be brought do a direct match with the previous one. When the same stimulation arrives to the right hemisphere, there is no learning being automatic or limited to
the incoming information, but a real spatial distribution process among intimately connected element starts. The right hemisphere associates with the ‘tree’ element all of its
fundamental components and associates with these other segments, integrating in this way the entire natural process.
Although the basic elaboration is separated and strictly autonomous, right and left hemisphere
constantly communicate through an anatomic connection bridge: the corpus callosum.
There are motor areas (Broca’s center and writing center) mainly active in the left hemisphere,
through which the right hemisphere’s stimulation must pass to express, through an articulate and comprehensible language, the concept of integration operating in the analysis of the
incoming inputs. To do this, the inside stimulation has to necessarily pass through the area of memorization of the model, modifying it through a energetic-biochemical (RNA, DNA)
that determines the break or the usage of the model.
In front of an incoming input, the learning function of the left hemisphere is based on the
binary code (same system used by electronic calculators):
- Or the datum is recognized as already memorized (obviously, in the case of a new datum it is directly memorized) and then it is identified with a recall datum
- Or it is in antithesis with the memorized one , as it does not present identically the same model of it, but offers a further structure model of the same element.
In this way the left hemisphere acts by selecting incoming data by exclusion: ‘yes’ or ‘no’, ‘true’ or ‘false’, there’s no third one that includes, by integrating them, two different aspects of reality.
Such a possibility is the fulcrum-basis of the right hemisphere, to which two different visions of the same concept a particular element may
be brought are synonym of a larger analysis space that allows to pick a larger number of particulars and, at the same time, offer a starting point for an associative study that
researches, if it wasn’t clearly visible, a relationship between the two aspects.
It is anyway important underlining that it would be a mistake reduce left and right hemisphere
respectively to a strong determinism and to a probabilism typical of two physical and structural models of matter and energy, as once again this would mean entering a concept in a block
scheme and again selecting it into models, i.e. observing it only through the left hemisphere.
If the input carries the information A=B, the left hemisphere learns A=B and identifies it with
a cultural model. This particular process is at the basis of all of today’s scientific knowledge, as all the branches of science, to study a phenomenon, hark back to a model which
characterize a certain number of parameters, sufficient to understand the phenomenon. The so-built construction of an enough satisfying model automatically morphs into a mental
model to be defended, difficult to be removed. The history of science is full of examples, and although it may look like a contradiction in terms, if not explained by cerebral physiology,
the field of cognitive learning that should theoretically be the most open to innovations and observation aiming to enlarge the knowledge, is often revealed to be dogmatically closed in
its prejudices.
A very deeper scientific analysis of the input is performed by the right hemisphere which splits
the equality A=B and determines the two elements A and B in a sequential succession, establishing that the verifying of the relationship A=B is impossible. This allows the
right hemisphere to preserve a spatial memorization freedom always open to any new stimulation, allowing it in a second moment a creative dynamic response. The right hemisphere doesn’t
even admit the possibility of a relationship A=A, as it is implied in the reality of A its uniqueness and individuality not reproducible in the equality.
The learning of a model by the left hemisphere and the analysis of the related parameters to an
element of the observable reality by the right hemisphere, follow a precise succession of nervous ways, being related to the differentiation of sensorial receptors and consequent areas of
association and elaboration. As for such a succession two kinds of coding are established: a biochemical and energetic coding based on the models in the left hemisphere, and an
energetic consequential spatial that personalizes the particulars, giving them a precise worth, in the right hemisphere.
The optical afferences characterize, in the left hemisphere, a formal model through which a
particular is identified which, to be associated in our mind to what we learnt in that particular, has to be represented visually in the same way. The optical afferences determine in the
right hemisphere the specification of the spatial-electromagnetic characteristics, through the integration of the contribution of the variable of every parameter.
The acoustic afferences carry into the left hemisphere a coding which is instantly identified with the code model that, in the language, will
be consequently associated every time the sensorial stimulations offer such information that, brought together in a global model, characterize that element. To the right hemisphere
the acoustic afferences determine an analysis of the conceptual reality of the linguistic code at the base of the emission spectrum that accompanies the same letter and to the same letter
(and every letter has an individuality of its own in the construction of a response, always preserving its conceptual meaning): definitely they associate to the structural reality
of the element the quantized impulses of an energetic code of a human size. The three aggregation statuses of the matter are a sign of as many energetic global statuses of a system, so
that a precise energy level corresponds to every status. Every element, even of a high density, owns a lot of sublimed which constitute the steam tension, often not
measurable.
The olfactory afferences in the left hemisphere constitute an aromatic model always associated
to that particular and determine it unambiguously. Such afferences bring, in the right hemisphere, to the chemical-energetic characterization of the structural part of the element to a
higher energetic global worth, i.e. the sublimed part. On this line such areas perform a first particular chemical analysis.
For the right hemisphere the analysis of taste afferences is added to the one above, while the
same afferences constitute a further model of taste for the left hemisphere: only that specific model will be associated to the element.
In the right hemisphere the taste area determines a specific chemical analysis of the element,
apart from the sublimed part, characterizing its acidity, the bond potentials and all of the other fundamental chemical characteristics. The synthesis of the olfactory and taste
afferences allows a complete chemical analysis of the element.
The fifth sensorial channel is the one of the tactile afferences which creates in the left
hemisphere a further model correlating the density of the element and its morphology in an unambiguous way and totally defines its stimulating tactile components corresponding to the
construction of the model (hard – soft – morphology; liquid – solid). In the right hemisphere the tactile afference determines the characteristics of density and completely
integrates the analysis of the element, so obtaining a complete spectrum of the characteristics which individuate it and, definitely, reflects the analysis performed by integrating
all of the energetic natural parameters.
Our sensorial instruments are a very powerful tool to study the reality around us and their
analysis spectrum corresponding to the physiology of the associative sensorial areas represents a fundamental example to understand which parameters are to be studied in that
phenomenon.
The right hemisphere integrates in its analysis all of the physical scientific parameters which
constitute the message, as it analyzes from an energetic, physical, chemical, biological point of view and synthesizes the results coming from it to elaborate a complete response on the
objective reality of the element. Such an objective reality is deformed when the right hemisphere transfers the message through the corpus callosum to the structural conformation of
the cultural models of the left hemispheres, which has to match, integrate and express, through its own codes, the objectivity of the right hemisphere, so making impossible the
transmission of the message in its initial integrity.
Left and right hemisphere pick two different aspects of the world around us. The first one
learns and memorizes an artificial reality, made by models and combinations of these which generate further models, and the coding which takes place within the hemisphere is in the same
way artificial. The second one analyzes and records the reality of the physical-natural world and the coding within this hemisphere reflects such a naturalness.
In the presence of a stimulation, the left hemisphere quickly emits a response, i.e. acts on
stimulus-response, while the right hemisphere, through a less quick process, emits a new then response due to how its physiological inner activity sets the result (always new,
corresponding to the news of the last interactive moment).
Although left and right hemisphere are such physiologically differenced aspects of our brain,
they are not in antithesis, on the contrary their physiology has to further be integrated through a higher contribution of interhemispherical interactions.
The same physiology gives us the example of a double synchronous activity of the vegetative
system, and other examples could be drawn not only from the human physiology, but from the whole physiology and anatomy of the natural world (to be considered as mechanisms of working and
integrated structures).
The first step to the knowledge of the reality of this ecosystem of ours is the knowledge of our
cerebral physiology and how it accomplishes the world within; to reach such a knowledge we have to perform on ourselves such an experimentation that the right hemisphere breaks the
schemes and cultural models, so to transfer to the left hemisphere a new cognitive methodology which doesn’t bring anymore to a repetitive culture, but a creative one, i.e. to a
conscious comprehension of the world around us and ourselves: this work doesn’t aim to create a new dominant hemisphere, but to search for a new integrative synthesis of the depolarities
of our brain.
In short, we can highlight which should physiologically be the path of the physical message
building the individual, in input, to our senses. It is received by the two hemispheres: if the preformed message has a possibility of integrative continuity with the new incoming
messages, it is built by the left hemisphere in such a way to allow the right hemisphere to enrich, measure and arrange the construction of the objective reality of the received message.
If the message is considered as ‘truth’, in the left hemisphere a neurological model will be formed which can not be integrated with other messages, not to be touched, and the
biological mechanism of defence will be activated every time there will be an attempt to alter such a message, both by the right hemisphere and the outside environment. In the first case
the right hemisphere is allowed to perform a creative action on the world around not identifiable in a preformed model, allowing the receiver of such an action to elaborate a new
message.
This physiological process accelerates the human evolution and allows every brain to be busy on
creative psychological constructions, giving it the pleasure to be protagonist of ideas and removing from it the need to aggress in order to protect models and messages which due to their
formation are closed to any possibility of integration of different stimulations not identifiable by the brain among the memorized models.
It’s up to us to be able to arrange the culture in such a way that any sensorial message coming
from any sense organ may run through the entire thick nervous net without the borders created by the repetitive cultural models.

 ....Teorie Scientifiche....
....Teorie Scientifiche....



FILOGENESI DELLA COSCIENZA
Discorso di compleanno del Prof. Michele Trimarchi Primo Novembre 2018